A peu près 85% des variations d’intelligence entre les individus et entre les populations sont des variations génétiques. Une personne intelligente aura un ensemble de versions alléliques favorables. Voici les arguments qui le démontrent sans l’ombre d’un doute.
- 1. Haut niveau d’héritabilité
- 2. Grande stabilité des Q.I raciaux quel que soit le milieu
- 3. Stabilité dans le temps
- 4. Les différences de Q.I sont reflétées dans les différences de volumes crâniens
- 5. Les temps d’isolation
- 6. Les études sur les métis
- 7. Les différences cérébrales qualitatives
- 8. Les études d’adoption
- 9. Différences raciales dans les temps de réaction
- 10. “Inspection time”
- 11. Admixture européenne chez les Afro-Américains
- 12. 76 traits musculo-squelettique, proximité avec homo erectus
- 13. L’intelligence s’inscrit dans un ensemble de traits évolutifs
- 14. Position dans l’évolution
- 15. Régression vers la moyenne
- 16. Inbreeding depression
- 17. Différences raciales à l’EEG
- 18. Spearman’s hypothesis
- 19. L’héritabilité augmente avec l’âge
- 20. Différences raciales dans les vitesses de maturation intellectuelle
- 21. Existence des différences raciales dans l’intelligence depuis 10 mille ans
- 22. Différences de fréquence des gènes de l’intelligence
- Conclusion
- Références

James Watson, prix Nobel 1962 pour la découverte de la structure de l’Adn.
1. Haut niveau d’héritabilité
Le haut niveau d’héritabilité trouvé chez les vrais jumeaux en Amérique, en Europe, au Japon et en Inde montre que l’intelligence est fortement déterminée par des facteurs génétiques.
- La corrélation entre vrais jumeaux élevés dans des milieux différents est de 0,85 à l’âge adulte.
- La corrélation intellectuelle entre deux individus de parents biologiques différents élevés dans le même milieu atteint 0 à l’âge adulte1.
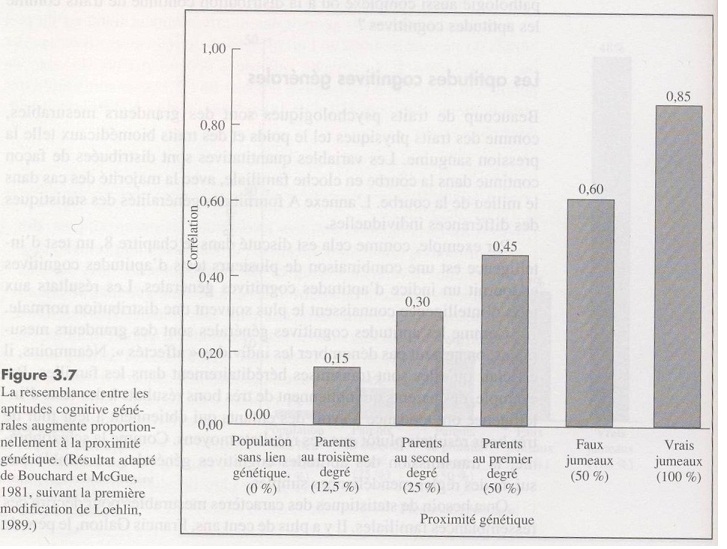
Tableau extrait de « Des gènes au comportement », Plomin, de Fries, Mc Clearn, Rutter, 1997.
→ Pointe clairement la causalité génétique de l’intelligence (trait polygénique)
A l’inverse, la corrélation intellectuelle entre des frères et sœurs adoptifs permet une estimation de l’importance du milieu. La corrélation est de 0 à l’âge adulte. Aussi surprenant que cela puisse paraître, le milieu de vie ou l’éducation n’a pas d’impact sur l’intelligence générale à l’âge adulte1.
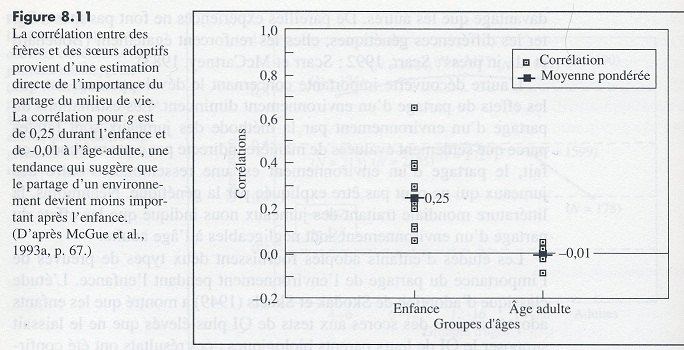
Tableau extrait de « Des gènes au comportement », Plomin, de Fries, Mc Clearn, Rutter, 1997.
Quelle est l’héritabilité de l’intelligence ?
→ Héritabilité de l’intelligence = part de la variance attribuable à des facteurs génétiques. Il y a plusieurs manières de l’estimer… pointant toute une héritabilité d’à peu près ~0,8 à l’âge adulte.
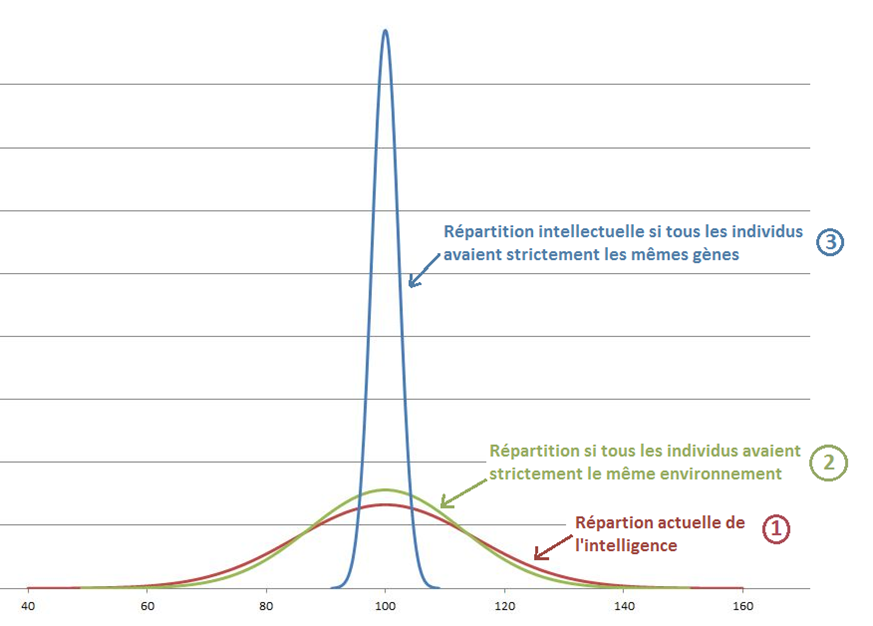
Illustration de la part génétique et de la part « environnementale » de l’intelligence2
Ceci signifie que si tout le monde était élevé dans un environnement identique, les différences intellectuelles entre individus seraient réduites à 80% des différences actuelles.
Il y a 3 manières de l’estimer… pointant toute une héritabilité > à 0,8 à l’âge adulte3.
- Etudes de jumeaux vrais élevés dans des milieux différents (Données résumées par Bouchard, 1993, p.58). Chez les adultes, les données sont basées sur 5 études résumées par Bouchard. La corrélation moyenne pondérée pour la taille des échantillons est de 0,75. Ce chiffre doit être corrigé pour la fiabilité des tests (correction de l’atténuation), (Bouchard, 1993, p.49; Machintosh, 1998). Cette correction augmente la corrélation à 0,83.
- Une seconde méthode pour estimer l’héritabilité est de comparer le degré de similitude entre des jumeaux identiques et des jumeaux non-identiques, de même sexe, élevés dans la même famille. La corrélation est de 0,88 pour des jumeaux identiques et de 0,51 pour des jumeaux non-identiques de même sexe. Après correction de ces chiffres pour la fiabilité des tests (correction d’atténuation), la corrélation corrigée devient 0,98 pour les jumeaux identiques et 0,56 pour les jumeaux de même sexe non identiques.
L’héritabilité peut être calculée par la formule de Falconer (1960) consistant à doubler la différence entre les corrélations de jumeaux identiques et non-identiques de même sexe. La différence entre les deux corrélations est de 0,42, doubler cette différence donne une héritabilité de 0,84. - Une troisième méthode pour estimer l’héritabilité est d’examiner la corrélation du Q.I entre enfants de parents biologiques différents adoptés et élevés dans les mêmes familles. Ceci permet d’estimer la magnitude de l’effet de l’environnement familial (« between family effect »). Le résumé de la littérature sur la question a été fait par Bouchard (1998) qui conclut que chez les adultes la corrélation est de 0,04, indiquant une héritabilité de 0,96.
Cependant, cette méthode sous-estime les effets environnementaux car elle ne prend pas en compte l’environnement prénatal et périnatal.
→ Les deux premières méthodes sont plus précises et donnent une estimation de 0,84 et 0,85 pour l’héritabilité de l’intelligence.
La conclusion d’une haute héritabilité pour l’intelligence implique qu’il y a des gènes qui déterminent l’intelligence (voir plus loin). Le premier fut découvert par Chorley et al. (1998). Il se situe sur le chromosome 6 et la possession d’un des allèles de ce gène contribue à hauteur de 4 points de Q.I.
2. Grande stabilité du Q.I des différentes races quel que soit le milieu4
→ appuie la causalité génétique. (Arabes d’Europe de première et seconde génération, immigration…)
Légère augmentation (84 à 88 pour les arabes… les gènes « tiennent l’amélioration en laisse4.2 »)
- Juifs Ashkénazes (110) (Amérique, Europe…)
- Asiatiques de l’est (105) (Asie, Europe, Amérique…)
- Européens (100) (Australie, Amérique, Europe…)
- Inuits (91)
- Métis Caucasien-Africain (90) (Europe, Amérique…)
- Asiatiques du sud-est (90) (Europe, Amérique, Asie)
- Amérindiens (87) (Amérique du nord et du sud)
- Nord Africains et Sud Asiatiques (86) (Moyen-Orient, Amérique, Europe…)
- Africains (80) (Europe, Amérique du nord et du sud, Afrique)
- Aborigènes d’Australie (62)
Cette hiérarchie reste rigoureusement inchangée et transparaît dans tous les pays multiraciaux4.1 dans quasiment tous les paramètres sociaux (salaires, niveaux socio-économique, niveau de crimes et délits, prévalence des personnes douées, prévalence de l’arriération mentale… voir « Hiérarchie intellectuelle à travers le monde »)
3. Stabilité dans le temps4.2
Pointe une nouvelle fois l’héritabilité élevée de l’intelligence.

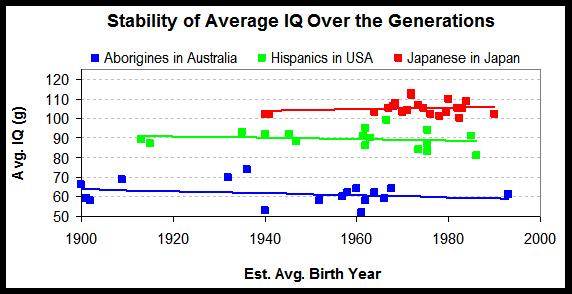
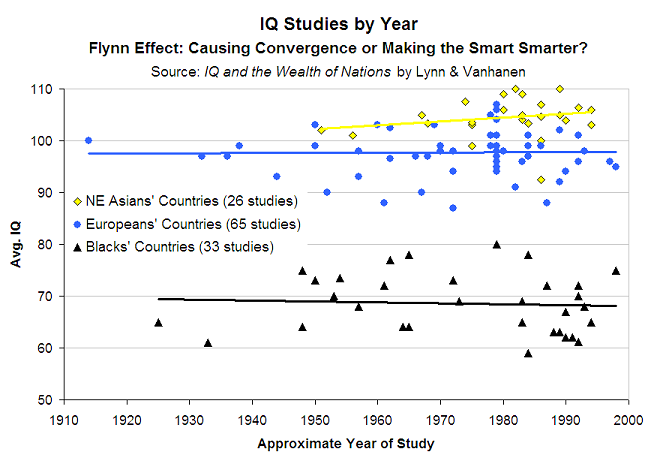
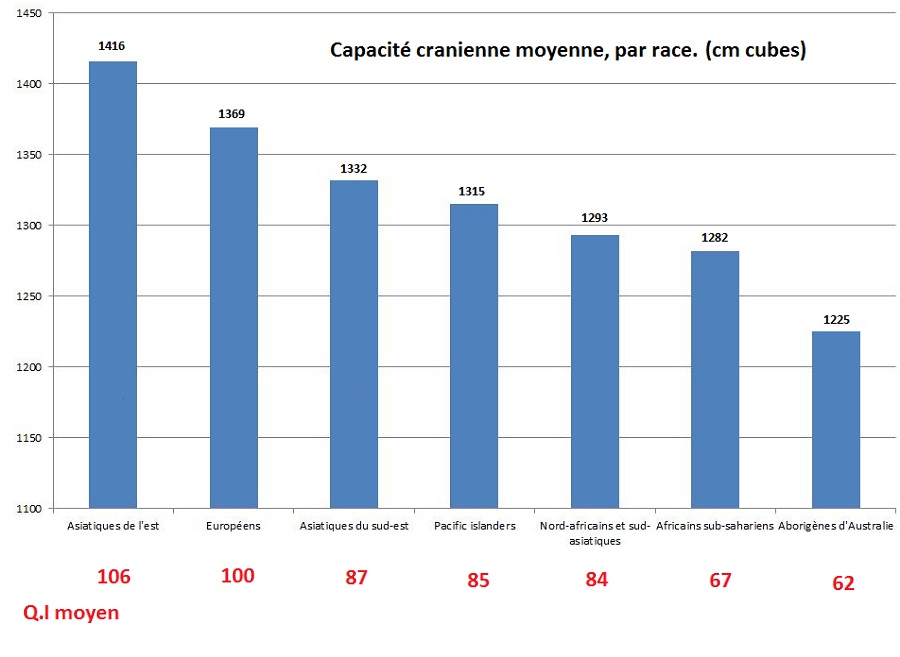
4. les différences de Q.I sont reflétées dans les différences de volume crânien5
Table 9.8. Race differences in brain site (cc) and intelligence
Race IQ Brain Size (cc) East Asians 105 1416 Europeans 100 1369 Southeast Asians 90 1332 Pacific Islanders 85 1317 South Asians and North Africans 84 1293 Africans 67 1282 Australian aborigenes 62 1225
Ces différences de volumes crâniens démontrent l’existence de facteurs génétiques, car l’héritabilité des volumes crâniens est de 0,9 et la corrélation entre l’intelligence et volume crânien est de 0,45.

Capacité cranienne et Q.I chez les Africains, les Européens et les Est-Asiatiques6.
5. Les temps d’isolation.
Les différentes races ont évolué indépendamment l’une de l’autre et dans des milieux différents pendant des dizaines de milliers d’années. Par exemple, les Africains ont vécu dans des milieux séparés des Européens pendant 100.000 ans, et les Européens ont vécu dans des milieux séparés des Asiatiques de l’est pendant 40.000 ans (Stringer, 1989; Cavalli-Sforza, 2000)
Quand deux populations évoluent isolément l’une de l’autre pendant une telle période il y a des différences qui apparaissent dans tous les domaines dans lesquels il y a des possibilités de variations génétiques. La position environnementaliste extrême, postulant qu’il n’y a pas de différences intellectuelles entre deux races, défie le principe général de l’évolution biologique et peut être vu comme impossible.
6. les études sur les métis
Le Q.I des « hybrides » est toujours intermédiaire entre celui des deux races parentales, de même que le volume crânien, qui est lui aussi intermédiaire entre celui des deux races parentales, ceci pointe la causalité génétique de l’intelligence.
6.1 Hybrides Africains/Européens7.
EUROPEANS HYBRIDS AFRICANS Location Age Test N IQ N IQ N IQ Reference 1 Brazil 10 SPM 735 95 718 81 223 71 Fernandez, 2001 2 Germany 5-13 WISC 1,099 100 170 94 – – Eyferth, 1961 3 South Africa 10-12 AAB 10,000 100 6,196 83 293 65 Fick, 1929 4 South Africa 13 GSAT 746 100 815 86 – – Claassen, 1990 5 South Africa 15 SPM 1,056 100 778 80 1,093 74 Owen, 1992 6 USA 17 WISC- R 16 102 55 94 17 85 Weinberg et al., 1992 7 USA Adult Otis – 100 284 91 176 87 Codwell, 1947 8 USA Adult Vocab 1,245 100 304 92 146 85 Lynn, 2002 9 USA Adult Vocab 10,315 100 116 97 4,271 89 Rowe, 2002 10 Canada 7-12 PP 100 – 46 93 46 78 Tanser, 1941
6.2 Hybrides Amérindiens/Européens8
Table 12.4. IQs of Native American-European Hybrids
Location Age Test Europeans Hybrids Amerinds Reference N IQ N IQ N IQ 1 Kansas Adult OTIS - 100 536 93 179 67 Hunter & Sommermeir, 1922 2 South Dakota 10-15 OTIS - 100 68 89 15 86 Fitzgerald & Ludeman, 1925 3 Mexico 7-10 SPM 155 98 571 94 194 83 Lynn et al., 2005
6.3 Hybrides Pacific-Islander/Européens9
Q.I des hybrides Européens-Pacific Islanders et Chinois-Pacific Islanders
Table 9.3. IQs of Europeans, Chinese, and Pacific Islander Hybrids
Group N IQ European 1000 100 Chinese 2,704 99 European-Hawallan 842 93 Chinese-Hawallan 751 91 Hawalian 621 81
6.4 Hybrides Est-Asiatiques/Européens10
Table 10.5. IQs and brain size (cc) of East Asian-European hybrids
East Asian Hybrid European African 1 Number 63 37 17,432 19,419 2 IQ-Raw 114 103 102 90 3 IQ-Adjusted 109 98 97 85 4 Brain size 1,170 1,155 1,150 1,134
6.5 Hybrides Aborigènes d’Australie/Européens11.

7. Les différences cérébrales qualitatives
Les différentes races n’ont pas un cerveau identique de taille variable, il y a des différences qualitatives entre les races.
Africains et Européens12:
- Le cortex des Africains est en moyenne moins circonvolué.
- Le cortex des Africains est en moyenne 15% plus fin que celui des Européens.
- Les Africains ont un lobe frontal et occipital plus petit et un lobe pariétal plus large.
- Les Africains ont une proportion de neurones pyramidaux moindre à celle des Européens.
Aborigènes d’Australie et Européens13:

D’après « Race », John R. Baker, professeur de biologie à Oxford13.
On peut identifier avec précision l’ascendance raciale de quelqu’un par IRM (Imagerie par Résonance Magnétique)22 ceci parce que le cortex cérébral forme des repliements (gyrus et sillons) spécifiques à chaque ascendance raciale23.
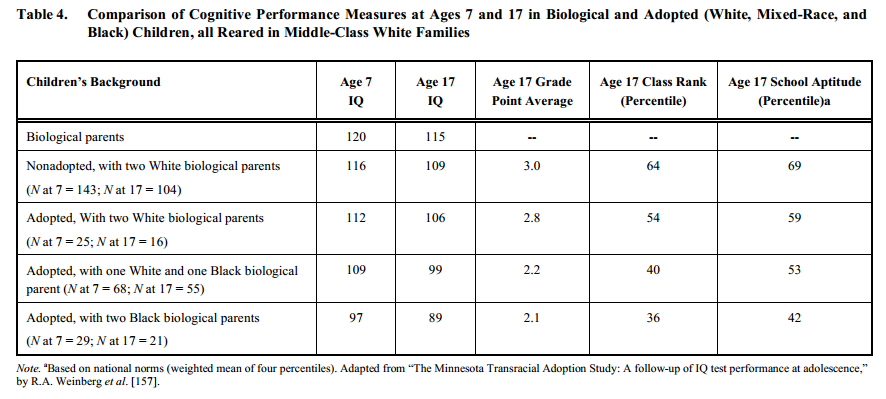
8. Les études d’adoption
Montrent clairement que le Q.I reste celui prédit par la race biologique quel que soit les parents adoptifs.
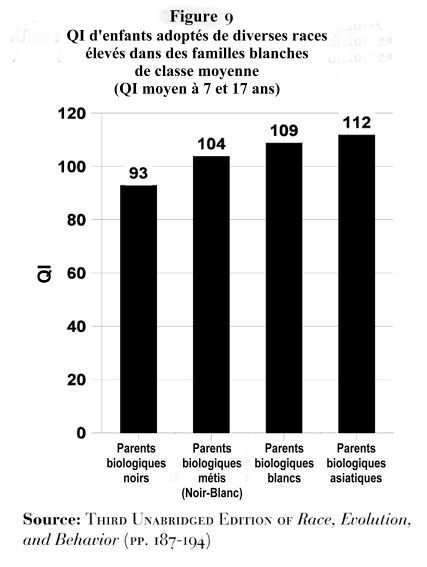
Q.I moyens d’enfants de parents biologiques Noirs, Métis, Blancs et Asiatiques (Est-Asiatiques), tous élevés dans des familles blanches de classe moyenne14.
Coréens adoptés par des belges: 106 de Q.I moyen.
Pour des enfants qui sont tous adoptés par des parents européens, il persiste une différence de 16 points de Q.I entre Africains et Européens, la même que celle trouvée en Amérique.
Elever des enfants noirs dans une famille blanche de classe moyenne n’a pas d’effet sur leur Q.I à 17ans.
L’éducation n’a pas plus d’effet sur les Est-Asiatiques élevés par des Blancs, leur Q.I moyen restant supérieur.
→ Facteurs génétiques.
9. « reaction time »
Il existe une différence significatives entre les Européens, les Africains et les Est-Asiatiques en termes de temps de réaction.
Le temps de réaction est corrélé au Q.I, car l’un comme l’autre sont des signes d’efficience du système nerveux central. Les caucasiens réagissent en moyenne plus vite à un stimulus.
L’individu testé est placé devant une petite lampe qui va s’allumer. Chaque fois que c’est le cas, il appuie simplement le plus rapidement possible sur le bouton placé devant lui.
C’est un signe de l’efficience du système nerveux puisque c’est en quelque sorte un traitement basique de l’information qui doit remonter jusqu’au cerveau puis redescendre pour appuyer sur le bouton. Les temps de réaction sont mesurés en millisecondes.
Reaction Times et EEGs chez des Africains, des Européens et des Est-Asiatiques4.3.
Test Africans Europeans East asians 1 IQ 68 100 106 2 RT-S 398 371 348 3 RT-C 1,950 1,220 - 4 EEG 534 506 -
Meilleure efficience du système nerveux central des hauts Q.I

10. « Inspection time »
« L’inspection time » ou temps d’inspection mesure la vitesse du traitement de l’information visuelle ou auditive. Les mesures « d’inspection time » sont corrélées à +0,7 avec le Q.I.
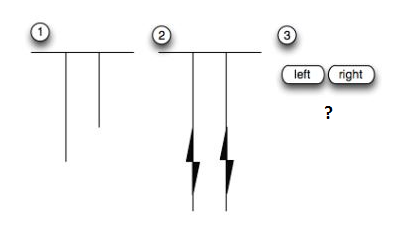
Dans ce genre de tests, deux barres de longueurs inégales apparaissent à l’écran pendant un certain temps (en millisecondes). On demande ensuite aux testés quelle était la barre la plus longue, celle de droite ou celle de gauche ?
Les hauts Q.I traitent l’information visuelle ou auditive plus rapidement. Ils ont des temps d’inspection (inspection time) plus petits.
On trouve une différence significative, en termes d’inspection time, entre les races.
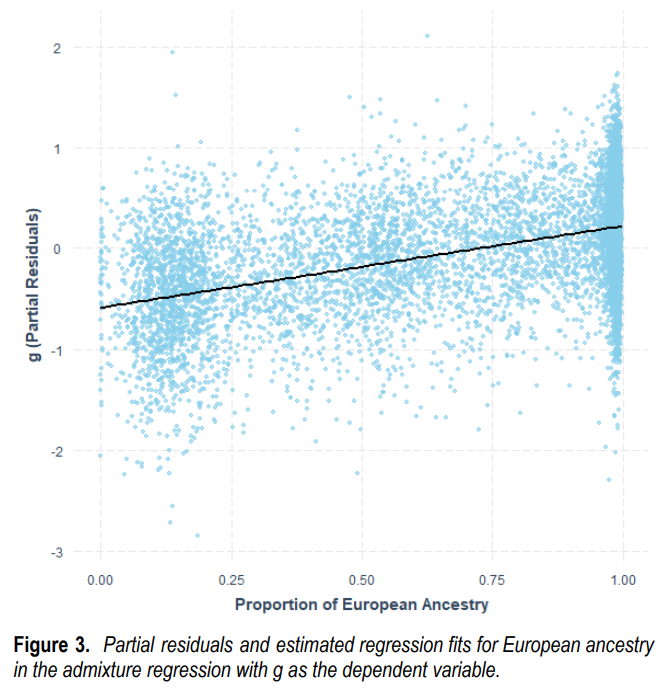
11. Admixture européenne chez les Afro-Américains.
Plus l’admixture européenne est importante chez les Afro-Américains, plus
(a) le poids cérébral moyen est élevé
(b) le QI moyen est élevé et plus
(c) le score polygénique d’intelligence est élevé
(a) Volume Cérébrale en fonction du % d’ascendance européenne chez les Afro-Américains (de 0 à 100%)
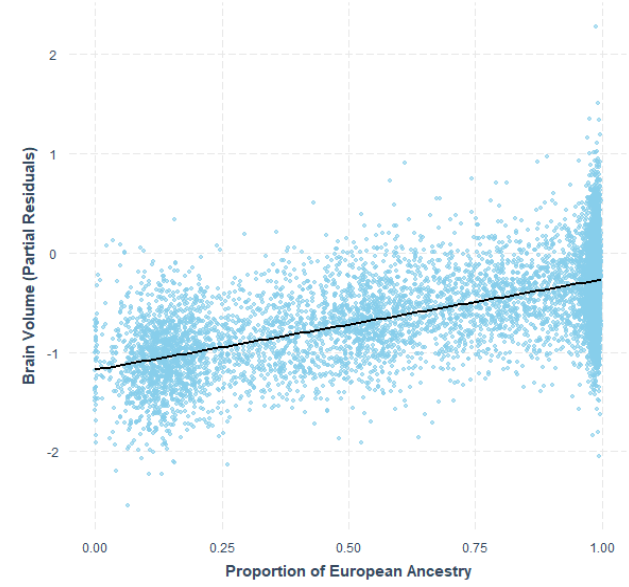
(b) QI moyen (g) en fonction du % de gènes européens chez les Afro-Américains (de 0 à 100%)
(Le QI moyen des Afro-Américains sans ascendance européenne est de 80; avec 25% d’ascendance européenne: 85 (Afro-Américains en moyenne); avec 50% d’ascendance européenne : 90; avec 75% d’ascendance européenne: 95)
(c) Scores polygéniques d’intelligence chez les Afro-Américains (ronds rouges) et chez les Européens (ronds mauves)
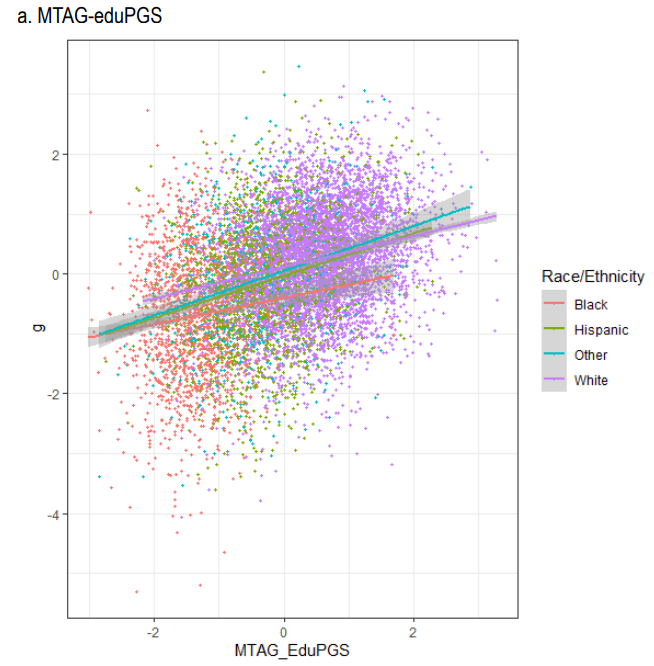
Références V. Shibaev et al. (2023) A Genetic Hypothesis for American Race/Ethnic Differences in Mean g: A Reply to Warne (2021) with Fifteen New Empirical Tests Using the ABCD Dataset
12. 76 traits musulo-squelettiques15
Les différences raciales de capacité crânienne sont corrélées à 76 traits musculo squelettiques identifiés dans des ouvrages standards d’anatomie évolutive comme étant systématiquement liés à une augmentation de la capacité crânienne et de l’intelligence chez les hominidés
Parmi ces différences, on retrouve :
-> Le diamètre transversal du pelvis : L’augmentation de la capacité crânienne et de l’intelligence a été de pair avec une augmentation du diamètre transversal du pelvis, pour permettre le passage du crâne à la naissance. Les africains ont un diamètre pelvien significativement plus petit que celui des Européens. (27,4 cm contre seulement 24,6 pour les africains). Les Est-Asiatiques ont un diamètre pelvien supérieur à celui des Européens.
-> En conséquence d’un pelvis plus large, le fémur, (l’os de la cuisse) qui s’insère au niveau du bassin, s’est incurvé puisqu’en dépit d’un bassin qui grandissait, espaçant les insérions fémorales et provoquant un angle plus large pour la sortie des deux fémurs, il fallait impérativement que le genou fasse une jonction correcte avec le péroné, provoquant une incurvation du fémur. Les Européens ont une incurvation fémorale significativement plus importante que celle des Africains et significativement moins importante que celle des Est-Asiatiques.
-> Tandis que l’intelligence et la capacité crânienne ont augmenté, les crânes sont devenus plus sphériques et profonds. Les Européens ont des cerveaux significativement plus sphériques, plus profonds et plus gros que les africains.
-> L’augmentation de la sphéricité a donc réduit les protubérances, notamment le processus mastoïde. Les Blancs ont un processus mastoïde significativement plus petit que les Noirs.
-> L’augmentation de la capacité crânienne s’est produite vers l’avant du crâne, il en a résulté une diminution du prognathisme et une augmentation de l’orthognathisme (face plus plate). Les Européens ont une face significativement moins prognathe et plus orthognathes que les Africains.
Progressive Changes in Brain Size and Musculo-Skeletal Traits in Seven Hominoid Populations (2004) J.P. Rushton , E.W. Rushton
En détail… (traduit de l’étude ci-dessus).
Troglodytes
(Chimpanzés)
Australopithèques
Homo
habilis
Homo
erectus
Homo
sapiens africains
Homo
sapiens européens
Homo
sapiens mongoloïdes
1. Intelligence générale (Q.I)
22
32
42
50
67-80
100
105
2. Capacité crânienne (cm cubes)
380 (1)
450 (2)
650 (3)
1000
(4)
1267
(5)
1346
(6)
1364
(7)
3. Quotient d’encéphalisation
3,01
(2)
2,79
(1)
3,38
(4)
3,34
(3)
6,38
(5)
6,50
(6)
6,95
(7)
Caractéristiques
du crane
4. Forme du crane
Le plus
étroit (1)
Etroit
(2)
Plus large que chez
l’australopithèque (3)
Plus large que chez homo
habilis (4)
Plus large que chez homo
erectus, le plus étroit des modernes (5)
Plus large que chez les
africains (6)
Le plus
large (7)
5. Longueur du crane
La plus
longue (1)
Longue
(2)
Plus courte que chez
l’Australopithèque (3)
Plus courte que chez homo
habilis (4)
Plus courte que chez homo
habilis, la plus longue des modernes (5)
Plus courte que chez les
africains, plus longue que chez les mongoloïdes (6)
La plus
courte (7)
6. Contour sagittal (circonférence
sagittale)
Valeur
la plus faible (1)
Faible valeur mais
légèrement supérieure à P. Troglodyte (2)
Valeur plus élevée que chez
les australopithèques (3)
Valeur plus élevée que celle
des homo habilis (4)
Valeur plus élevée que chez homo
erectus, valeur la plus faible des modernes, dépression post-bregmale (5)
Plus élevée que chez les
africains, moins que chez les mongoloïdes (6)
La plus
élevée (7)
7. Proéminence de l’os nasal
Plat
(2)
Plat
(2)
Plat
(2)
Légère
saillie (5)
Légère
saillie (5)
Le plus
saillant (7)
Légère
saillie (5)
8. Prognathisme facial
Le plus
prognathe (1)
Très
prognathe (2)
Moins prognathe que chez les
australopithèques (3)
Moins
prognathe qu’homo habilis (4)
Moins prognathe qu’homo
erectus, le plus prognathe des modernes (5)
Moins prognathe que les
africains, plus que les mongoloïdes (6)
Orthognathes
(7)
9. Largeur bi-zygomatique
La plus
large (1)
Large
(2)
Légèrement
rétrécie (3)
Plus étroit que chez homo habilis
(4)
Plus étroit que chez homo
erectus, la plus large des modernes (5)
Plus étroit que chez les
africains, plus large que chez les asiatiques (6)
La plus
étroite (7)
10. Forme du palais
En
forme de U (1)
Légèrement moins en forme de
U (2)
Presque en forme de V (3)
Plus en forme de V, presque
parabolique (4)
Le moins parabolique des
modernes (5)
Parabolique
(6)
Le plus
parabolique (7)
11. Forme de l’arcade sourcilière
Non archée et la plus large
(1)
Non
archée et large (2)
Archée et légèrement plus
petite (3)
Plus petite, archée et
arrondie avec dépression glabellaire (4)
Présence d’une dépression
glabellaire, la plus large des modernes (5)
Petite,
lisse et archée (6)
Archée et la plus petite (7)
12. Processus mastoïde
Le plus
large (1)
Large
(2)
Plus petit que chez les
australopithèques (3)
Plus petit que chez les homo
habilis (4)
Le plus large des modernes,
deux têtes (5)
Petit
et pointu (6)
Le plus
petit, trapu (7)
13. Position du neurocrâne
Derrière
la face (1,5)
Derrière
la face (1,5)
Plus haut mais toujours
derrière la face (3)
Plus haut que chez les homo
habilis mais toujours derrière la face (4)
Plus haut que chez les homo
erectus, mais le plus postérieur des modernes (5)
Sur le dessus de la face (6)
Le plus au-dessus de la face
(7)
14. Circularité du foramen temporal
Long et
étroit, ovale (1)
Court et étroit, mais pas
tout à fait circulaire (2)
-
Court
et étroit, circulaire (3)
-
-
-
15. Rotation faciale
Absente
(1)
Absente
à légère (2)
Légère
(3)
Présente
et plus proéminente (4)
-
-
-
16. Position antérieure
du foramen magnum et condyles occipitaux
Orienté de manière
postérieure, derrière la ligne bi-tympanique (1)
Plus antérieur que chez les
troglodytes, moins que chez les homo habilis (2)
Plus antérieur que chez les
australopithèques (3)
Proche de la position d’homo
sapiens (4)
-
-
-
17. Courbure de la suture squamozal
Toute
droite et basse (1,5)
Toute
droite et basse (1,5)
Incurvée
et basse (3)
Plus incurvée et plus haute
que chez homo habilis (4)
-
-
-
18. Orientation des condyles occipitaux
Vertical
(1)
Horizontal
(3)
Horizontal
(3)
Horizontal
(3)
-
-
-
19. Taille du muscle masséter
Large
(1,5)
Large
(1,5)
Plus petit que chez les
australopithèques (3)
Plus petit que chez homo
habilis (4)
-
-
-
20. Taille des constrictions post orbitales.
La plus
large (1)
Légèrement plus petite que
chez les troglodytes (2)
Légèrement plus petite que
chez les australopithèques (3)
Plus petite que chez homo
habilis (4)
Plus petite que chez homo
erectus, la plus large des modernes (5)
Plus large que chez les
africains, moins que chez les asiatiques (6)
La plus
petite (7)
Caractéristiques des
dents et de la mandibule
21. Forme des incisives
Plat
(2,5)
Plat
(2,5)
Plat
(2,5)
Plat
(2,5)
Rarement en forme de pelle
(5)
Plus souvent en forme de
pelle (6)
Souvent en forme de pelle
(7)
22. Nombre de dents
32 (3)
32 (3)
32 (3)
32 (3)
32 (3)
30-32
(6)
28-32
(7)
23. Taille des molaires
Plus petites que chez les
australopithèques (2)
Les
plus larges (1)
Plus petites que chez P.
Troglodytes (3)
Plus petites que chez homo
habilis (4)
Plus petites que chez l’homo
erectus, les plus larges des modernes (5)
Plus petites que chez les
africains (6)
Les
plus petites (7)
24. Orthognathisme de la mandibule
Longue,
basse, prognathe (1)
Légèrement plus courte et
plus haute, mais toujours prognathe (2)
Plus courte et plus haute
que chez les australopithèques (3)
Courte
et haute (4)
La plus longue et la plus
basse des modernes (5)
Taille
et hauteur intermédiaire (6)
Courte
et haute (orthognathe) (7)
25. Forme de la mandibule.
En
forme de U (1)
Plus proche de la forme d’un
V (2)
Presque en forme de V (3,5)
Presque en forme de V
(3,5)
La moins en forme de V des
modernes (5)
Moins en forme de V que chez
les asiatiques (6)
Le plus en forme de V (7)
26. Largeur entre les condyles mandibulaires
Le plus proche l’une de
l’autre (1)
Plus éloignées que chez le
troglodyte, plus proches que chez l’homo habilis (2)
Plus éloignées que chez les
australopithèques (3)
Plus éloignées que chez
l’homo habilis (4)
Plus éloignées que chez
l’homo erectus, le plus proche des modernes (5)
Plus large que chez les
africains, plus proches que chez les asiatiques (6)
Le plus
éloignée (7)
27. Proéminence du menton
Absente
(2)
Absente
(2)
Absente
(2)
Absente
(2)
Réduite
(4)
Proéminente
(7)
Modérée
(6)
28. Rapport entre la taille des condyles et la taille des
processus coronoïdes.
Plus
petits condyles (1)
Condyles un peu plus grands
(2)
Rapport
égal (3,5)
Rapport
égal (3,5)
-
-
-
29. Incisure mandibulaire
Peu
profonde (1,5)
Peu
profonde (1,5)
Profonde
(3,5)
Profonde
(3,5)
-
-
-
30. Etroitesse du ramus ascendant
Le plus
large, antéropostérieur (1)
Large,
antéropostérieur (2,5)
Large,
antéropostérieur (2,5)
Etroit,
antéropostérieurs (4)
-
-
-
31. Longueur de la racine des dents
Les
plus longues (1)
Longue
(2)
Plus courtes que chez les
australopithèques (3)
Plus courtes que chez homo
habilis (4)
Plus courte que chez homo
erectus (5)
Les
plus courtes (6,5)
Les
plus courtes (6,5)
Caractéristiques
du cou
32. Taille de la région du cou
La plus
haute (1)
Haute
(2)
Basse
(3)
La plus
basse (4)
33. Forme du cou
Large
et trapue (1,5)
Large
et trapue (1,5)
-
-
Plus
étroit (5)
Etroit,
courbé et long (6)
-
34. Masse des muscles du cou
La plus
importante (1)
Réduite par rapport à P.
Troglodyte (2)
Réduite par rapport à
Australopithèque (3)
Réduite,
proche des modernes (4)
Réduite, la plus large des
modernes (5)
Petite
(6)
La plus
petite (7)
35. Complexité des muscles
Complexe (nombreux flancs et
corps) (1)
Moins complexe que P.
Troglodyte (2)
Similaire
aux australopithèques (3)
Simple (moins de flancs et
de corps)
-
-
-
36. Proximité des muscles rectus capitis
Cote à
cote (1)
Ecartés
(2)
Encore
plus écarté (3,5)
Similaire
à H. Habilis (3,5)
-
-
-
37. Taille de la crête nucale et des marquages osseux
Présents
et larges (1)
Présents chez la plupart,
réduit chez certains (2)
Marquage
moins marqués (3)
Marquage
encore moins marqué (4)
Marquage
très peu marqué (5)
Lisse à
absente (6)
Absente
(7)
Caractéristiques
des vertèbres
38. Plus longue apophyse épineuse
5ème ou
6ème (1)
Le plus fréquemment la 6ème (2)
-
7ème,
comme les modernes (3)
-
-
-
39. Taille des apophyses épineuses
La plus
longue (1)
Longue
(2)
-
Courte, mais légèrement plus
longue que chez les modernes (3)
-
-
-
40. Convexité de la partie supérieure de l’atlas
Concave
(1)
Moins
concave (2)
-
Plat
(3)
-
-
-
Caractéristiques
du pelvis
41. Diamètre transversal du pelvis (mm)
98 (1)
99.1
(2)
-
Dans le range des homo
sapiens (3,5)
122
(3,5)
132 (5)
-
42. Taille du diamètre antéro-postérieur (mm)
85
(1,5)
85
(1,5)
-
Dans le range des homo
sapiens (3,5)
103
(3,5)
118 (5)
-
43. Largeur bi-iliaque (mm)
132 (1)
200 (2)
-
-
250 (3)
270 (5)
252 (4)
44. Forme de la lame iliaque
Grande
et étroite (1)
Courte
et large (2)
-
Courte
et large (3)
-
-
-
45. Entaille sciatique
Absente,
peu développée (1)
Présente,
bien développée (2)
-
Dimorphisme sexuel, bien
développée comme chez les modernes (3)
-
-
-
46. Convexité de la branche descendante du pubis
Convexe
(1)
Droite
(2,5)
Droite
(2,5)
-
-
-
-
47. Taille de l’acétabulum (mm)
0,82
(1)
0,78-0,83
(2)
-
Proche
des modernes, 0,9(3)
-
-
-
48. Taille du sacrum
La plus
petite (1)
Plus
large (2)
-
Large,
comme les modernes (3)
-
-
-
Caractéristiques
des membres supérieurs
49. Orientation de la cavité glénoïde
Orientée
vers le crane (1)
Moins orientée vers le crane
que chez P. Troglodyte (2)
-
Orientée latéralement, comme
chez les modernes (3)
-
-
-
50. Taille du bras
Long
(1)
Long
(2)
-
Range
des modernes (3)
Le plus long des modernes
(4)
Plus long que les
asiatiques, plus petit que chez les africains (5)
Le plus
petit (6)
51. Capacité de l’extension du coude
Hyper
extension possible (1,5)
Hyper
extension possible (1,5)
Légère aptitude à l’hyper
extension (3)
-
-
-
-
Caractéristiques
des membres inférieurs
52. Ligne inter trochantérienne
Absente
(1)
Présente chez les grands
spécimens uniquement (2)
-
Présent
(3)
-
-
-
53. Taille de la tête fémorale
La plus
petite (1)
Intermédiaire entre les
singes et les humains (2)
Plus large que chez les
australopithèques (3)
Large
(4)
La plus petite des modernes
(5)
Intermédiaire entre les
africains et les asiatiques (6)
La plus
large (7)
54. Profile du condyle fémoral latéral
Circulaire
(1)
Circulaire chez les petits
spécimens mais plus ovale chez les grands spécimens (2)
Proche d’australopithèque
mais moins circulaire (3)
Presque
comme les modernes (4)
Le plus circulaire des
modernes (5)
Elliptique
(6)
Ovale
(7)
55. Symétrie des condyles fémoraux
Asymétrique
(1)
Asymétrique dans quelques
petits spécimens, plus symétrique dans d’autres spécimens (2,5)
Comme
l’australopithèque (2,5)
Symétrique
(4)
-
-
-
56. Largeur fémorale bi-condylaire (mm)
La plus
petite (1,5)
La plus
petite (1,5)
Légèrement
plus large (3)
Plus large, proche des
modernes (4)
79,50
83,05
-
57. Axe de l’angle condylien
Le plus
large (1,5)
Pareil
aux singes (1,5)
Légèrement
réduit (3)
Plus petit que chez H.
Habilis (4)
-
-
-
58. Minceur d’une section du col du fémur
Epaisseur corticale tout le
long, rond (1)
Os
cortical compressé (2,5)
Os
cortical compressé (2,5)
Comme les modernes, mais os
cortical légèrement plus épais (4)
-
-
-
59. Profondeur de la fosse trochantérienne fémorale
Profonde
(1,5)
Profonde
(1,5)
-
Moins
profonde (3)
-
-
-
60. Pilastre fémoral
Pas de
pilastre (2)
Pas de
pilastre (2)
Pas de
pilastre (2)
Pas de pilastre, parfois
petit pilastre (4)
Petit
pilastre (5)
Large
pilastre (6)
Le plus large des pilastres
(7)
61. Indice de courbure de l’axe fémoral
77,5
(droite) (2)
80 (3)
-
-
76,6
(1)
97,0
(4)
102,2
(5)
62. Taille du plateau tibial
Le plus
petit (1)
Légèrement plus large que
chez P. Troglodyte (2)
-
-
Le plus petit des modernes
(3)
Intermédiaire entre les
africains et les asiatiques (4)
Le plus
large (5)
63. Platitude du plateau tibial
Le plus
courbé (1,5)
Le plus
courbé (1,5)
-
-
Courbé
(3)
Plat
(4,5)
Plat
(4,5)
64. Concavité du condyle tibial latéral
Convexe
(1,5)
Convexe
(1,5)
Légèrement moins convexe que
chez les australopithèques (3)
-
Plat
(4)
Varie
(5)
Concave
(6)
65. Congruence du genou
La plus
basse (1)
Basse
(2)
Modérée
(3)
Haute
(4)
-
-
-
66. Angle du plateau tibial
Le plus
important (2)
Important
(2)
Plus
petit (3)
-
Le plus important de
modernes (4)
Plus
petit (5)
-
67. Taille du condyle tibial
Le plus petit et le plus
étroit (1)
Petit
et étroit (2,5)
Petit
et étroit (2,5)
Plus large, proche des
modernes (4)
Plus large qu’H. erectus, le
plus petit des modernes (5)
Plus large que les africains
(6)
Le plus
large (7)
68. Taille de la tête tibiale distale
Petit
(2)
Petit
(2)
Petit
(2)
Légèrement
élargie (4)
Plus
large qu’H. erectus, 47,07 (5)
Le plus
large, 53,23 (6)
-
69. Tranchant du bord antérieur du tibia
Rond
(2)
Rond
(2)
Rond
(2)
-
arrondie
(4)
Tranchant
(5,5)
Tranchant
(5,5)
Proportion
du corps
70. Taille (m)
Moins
de 1m (1,5)
Moins
de 1m (1,5)
1-1,5m
(3)
1,3-1,5
(4)
1,42-1,69
(5,5)
1,64-1,74
(7)
1,59-1,68
(5,5)
71. Indice inter- membral (Membres supérieurs comparés à la
taille des membres inférieurs)
108,7
(1)
85 (2)
86,5
(3)
Homo
sapiens range (4)
70,3
(7)
70,5
(5)
71,1
(6)
72. Indice brachial
80,1
(1)
-
-
-
78,5
(2)
75,5
(4)
76,5
(3)
73. Indice crural
101,9
(1)
-
-
-
86,2
(2)
83,3
(4)
86,5
(3)
74. Taille du bras en % de la taille du corps
La plus
élevée (1)
Elevée
(2,5)
Elevée
(2,5)
Comme
les hommes modernes (4,5)
45,76
(4,5)
44,54
(6)
75. % du poids du corps des membres supérieurs
Le plus
important, 15,8 (1)
Important,
12 (2)
Important
(3)
Moindre,
comme les modernes (4)
-
-
-
76. Taille de la jambe en % de la taille du corps
Le plus
petit (1)
Petit
(2,5)
Petit
(2,5)
Comme
les hommes modernes (4)
56,42
(6)
54,98
(5)
-
77. % du poids du corps des membres inférieurs
24,2
(1)
28 (2)
-
30 (3)
-
-
-
Maturation
Durée
gestationnelle (semaines)
La plus
courte (1)
(2)
(3)
(4)
39 (5)
40 (6)
Légèrement supérieure à
celle des européens (7)
Age de la puberté (male)
Le plus
tôt (1)
(2)
(3)
(4)
(5)
(6)
Le plus
tardif (7)
Age de la puberté
(femelle)
Le plus
tôt
(2)
(3)
(4)
(5)
(6)
Le plus
tardif (7)
Age
de la ménarche
-
-
-
-
Plus
tôt (1)
Intermédiaire
(2)
Plus
tardif (3)
Age
de la ménopause
-
-
-
-
Plus
tôt (1)
Intermédiaire
(2)
Plus
tardif (7)
Développement
moteur
Le plus
rapide (1)
(2)
(3)
(4)
(5)
(6)
Le plus
lent (7)
Développement
squelettique
-
-
-
-
Le plus
rapide (1)
Intermédiaire
(2)
Le
moins rapide (3)
Fermeture
des sutures crâniennes
-
-
-
-
Le plus
tôt (1)
Intermédiaire
(2)
Le plus
tard (3)
Développement
dentaire
-
-
-
-
Le plus
rapide (1)
Intermédiaire
(2)
Le
moins rapide (3)
Eruption des dents de
sagesse
-
-
-
-
1-2 ans
plus tôt (1)
intermédiaire
(2)
1-2 ans
plus tard (3)
Age de la marche (mois)
-
-
-
-
11 (1)
12 (2)
13 (3)
Fréquence de double
ovulation (sur 1000)
-
-
-
-
>16
(1)
8 (2)
<4
(3)
Triplets
(par million)
-
-
-
-
1700
(1)
100 (2)
10 (3)
Quadruplet
(par million)
-
-
-
-
0 (1)
1 (2)
60 (3)
Longévité
-
-
-
-
Moindre
(1)
Intermédiaire
(2)
La plus
importante (3)
–
13. L’intelligence s’inscrit dans un ensemble de traits évolutifs
Les Asiatiques de l’est et les Africains sont aux deux extrémités d’un continuum avec les Européens dans une position intermédiaire, non seulement sur les scores moyens des tests cognitifs et les mesures de la taille du cerveau, mais aussi sur 60 variables incluant la vitesse de maturation, la personnalité, la reproduction et l’organisation sociale. Il semble improbable que des facteurs sociaux puissent produire ce modèle cohérent sur un ensemble de comportements si divers. Ceci montre que l’intelligence fait partie d’un processus évolutif plus large trouvant ses origines dans l’évolution.

Tiré de « Rushton’s contributions to the study of mental ability » Arthur R. Jensen, Personality and Individual Differences, 2012.
Les Africains sont en fait plus r-sélectionné que les Européens, qui sont plus r-sélectionnés que les Est-Asiatiques (gradient zoologique r/K). Pour plus de détails voir la perspective comparative de Rushton.
14. Position dans l’évolution
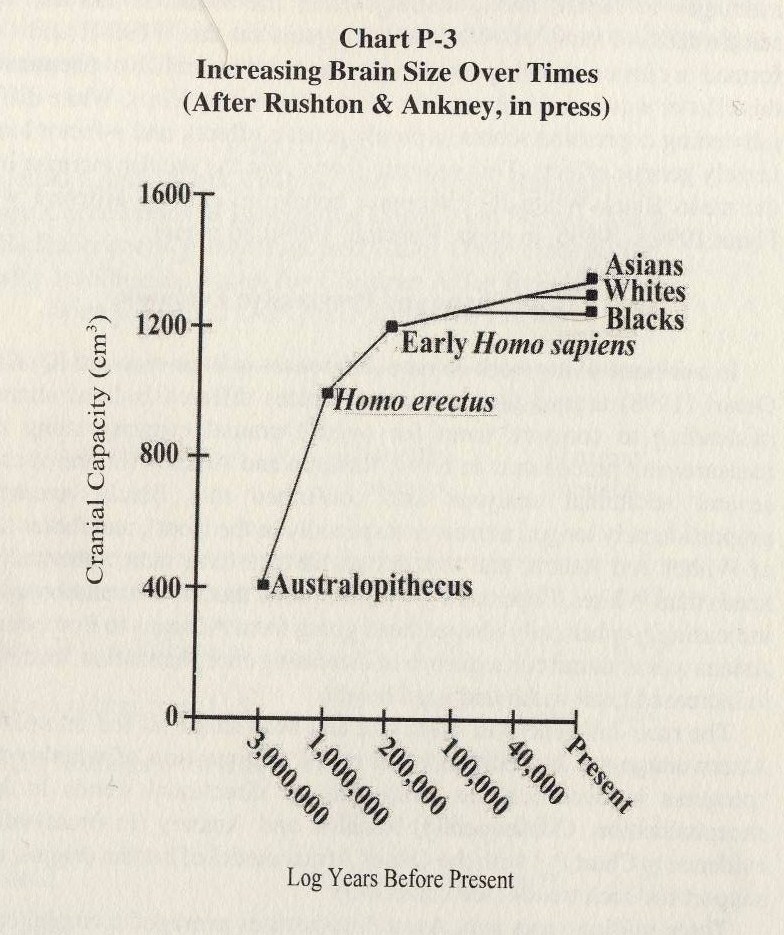
15. Régression vers la moyenne
Régression vers la moyenne pour la taille.
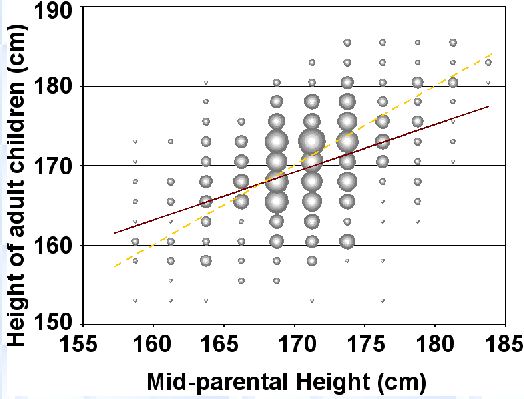
Régression vers la moyenne pour l’intelligence
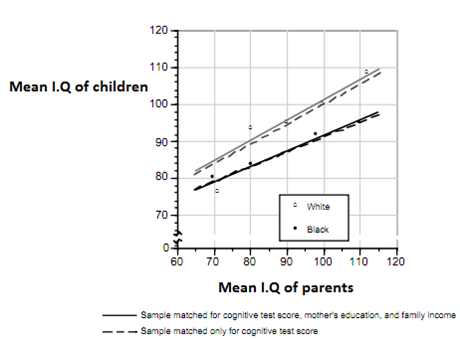
Comment comprendre le phénomène de régression vers la moyenne ?
Imaginez deux haricots de même taille: Le haricot A et le haricot B.
Le haricot A provient d’une race (=variété) de grands haricots. Le haricot B provient d’une race de petits haricots.
On plante les graines du haricot A et du haricot B. Qu’observe-t-on dans la descendance ?
La descendance du haricot A va régresser vers sa moyenne raciale plus élevée en taille: les haricots seront plus grands.
La descendance du haricot B va régresser vers la moyenne raciale de B, à savoir une plus petite taille.
Ce phénomène se retrouve très exactement en termes de Q.I et d’intelligence: prenez deux couples ayant un Q.I moyen de 110, un couple africain et un couple européen.
Les enfants du couple européen vont régresser vers le moyenne intellectuelle européenne de 100: ils auront en moyenne 105 de Q.I. Les enfants du couple africain Vont régresser vers la moyenne intellectuelle africaine de 80: ils auront en moyenne 95 de Q.I.
Ce phénomène de régression vers la moyenne se remarque également chez les bas Q.I: un enfant d’un couple européen ayant un Q.I moyen de 75 aura 87,5 de Q.I (régression vers la moyenne de 100), alors qu’un enfant d’un couple africain ayant en moyenne 75 de Q.I aura en moyenne 77,5 de Q.I (régression vers la moyenne de 80).
→ Signature de la moyenne génotypique européenne à 100 et de la moyenne génotypique africaine à 80.
Régression vers la moyenne pour l’intelligence
→ Signature de la causalité génétique de l’intelligence
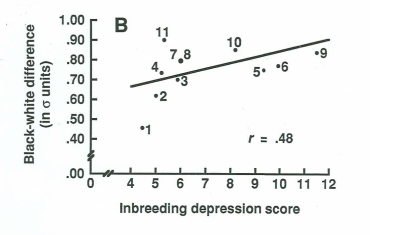
16. Inbreeding depression
- Diminution du « fitness » dans la descendance du croisement d’individus (ou d’animaux… l’homme est un animal) génétiquement trop proches
- Déficit de 7 points de Q.I chez les descendants de croisements entre cousins de première génération
→ Pointe la causalité génétique de l’intelligence.
17. Différences raciales à l’EEG16
Comme expliqué précédemment, les potentiels évoqués des individus plus intelligents ont un « score » plus petit: La fréquence des ondes alpha est plus importante.
En d’autres termes, la transmission de l’information est plus rapide chez les gens brillants.
On trouve une différence significative, en termes de score à l’EEG, entre les races.
Table 4.10. EEGs of Africans and Europeans
Test Africans in Africa Europeans in Europe 1 IQ 68 105 2 EEG 534 506
18. Spearman’s hypothesis
La magnitude de la différence intellectuelle entre deux races, observée dans les tests d’habileté cognitive, est proportionnelle à sa saturation en g15.
Exemple de la différence entre Européens et Afro-Américains. La différence culmine à 1 SD soit 15 points de Q.I.
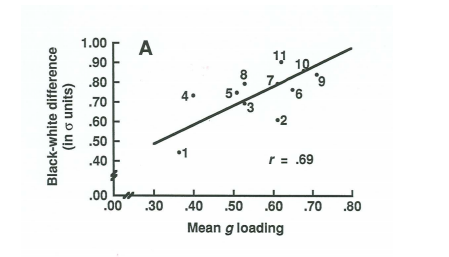
Autrement dit, plus un test sera saturé en g (plus il impliquera une complexité cognitive), plus la différence sera marquée.
→ Une différence de g est principalement responsable des différences intellectuelles raciales observées.
http://en.wikipedia.org/wiki/Spearman’s_hypothesis
19. L’héritabilité de l’intelligence augmente avec l’âge
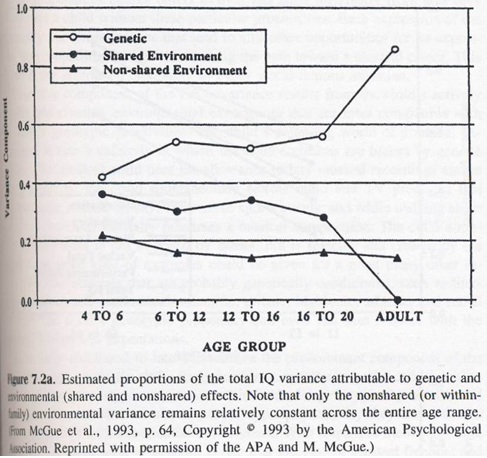
→ Ceci pointe la causalité génétique de l’intelligence.
Si l’environnement avait un impact, son importance devrait augmenter avec l’âge, or c’est le contraire qui se produit.
L’environnement non partagé cible essentiellement l’environnement in utero.
20. Différences raciales dans les vitesses de maturation intellectuelle
→ (1) Développement intellectuel le plus rapide chez les aborigènes d’Australie et intelligence finale la plus basse (62 de Q.I moyen)
→ (2) Développement intellectuel chez les africains un peu moins rapide que chez les aborigènes et intelligence finale un peu plus haute (71 de Q.I moyen)
→ (3) Développement intellectuel chez les Européens moins rapide que chez les africains et intelligence finale plus élevée (100 de Q.I moyen)
→ (4) Développement intellectuel le moins rapide chez les Est-Asiatiques et intelligence finale la plus élevée (105 de Q.I moyen)
Les Africains sont en avance sur les Européens dans leur développement intellectuel et moteur jusqu’à environs 30 mois (2,5 ans) puis ils sont rattrappés et dépassés.
Table 14.2. Differences between the means of South African Black and American White infants on the Bayley Scales of Infant Development, expressed as ds
AGE (mois): 6 12 21 30 Développement mental: 0,47 0,64 0 -0,01 Développement moteur: 0,94 0,24 0,06 -0,01
→ Les Aborigènes d’Australie ont aussi un faible Q.I (62) et ils montrent pareillement une précocité au stade de nourrisson. Ils sont plus avancés que les Caucasiens dans le contrôle du cou, du dos et des jambes (Freedman, 1974).
→ Développement intellectuel plus lent chez les Est-Asiatiques et intelligence finale plus élevée
Les enfants Européens sont en avance sur les Est-Asiatiques (pour le développement intellectuel et moteur) jusqu’à environs 8 ans.
Table 14.4. Norms for Taiwan for the Bayley Scales of Infant Development
AGE (mois): 6 12 18 24 Développement mental: -0.88 -0.82 -0.48 -0.23 Développement moteur: -1.06 -0.97 -0.74 -0.27
Table 14.3. Norms for Japan for the McCarthy Scales of Children’s Abilities
AGE (années): 2 3 4 5 6-8 Adultes Q.I: 94,4 96,7 97,1 97,9 99,1 105
Le principe bien connu de la biologie de l’évolution veut que les nourrissons d’espèces plus développées aient une enfance plus longue au cours de laquelle ils dépendent de leurs mères. Dès que les bébés reptiles éclosent de leurs œufs, ils peuvent se déplacer et se débrouiller par eux-mêmes, alors que les singes ont quelques années de dépendance maternelle. Parmi les primates, les plus primitifs sont les lémuriens, qui ont 2 ans de dépendance maternelle; les macaques sont un peu plus développés et ont 4 ans de prise en charge par la mère; les chimpanzés sont encore plus développés et exigent environ 8 ans de dépendance maternelle; et les humains sont les plus développés et ont environ 14 ans de dépendance maternelle (Lovejoy, 1981, Rushton, 2000, p.205).
J.P.Rushton a montré que ce principe s’étend aux trois principales races d’homo sapiens: les Est-Asiatiques ont un développement plus lent, une plus longue période de dépendance, et l’intelligence finale la plus élevée; Les Caucasiens (Européens, Sud-Asiatiques et Nord-Africains collectivement) mûrissent plus rapidement, tandis que les Africains sub-sahariens se développent le plus rapidement, ont la plus courte période de dépendance maternelle, et l’intelligence finale la plus basse. Ces différences sont présentes dans le développement physique, moteur et mental. Concernant le développement physique, les africains ont une maturité squelettique plus importante à la naissance, un développement dentaire plus rapide dans l’enfance et un développement sexuel plus rapide avec une adolescence plus précoce (mesurée par l’apparition des seins chez les filles et par le développement des organes génitaux chez les garçons). La théorie de Rushton a été confirmée par l’anthropologue japonais Kunihiko Kimura (Eiben, 1998).
Ces différences de vitesses de maturation intellectuelle signent à nouveau la causalité génétique des différences raciales d’intelligence, qui s’inscrivent dans un ensemble évolutif.

21. Existence des différences raciales dans l’intelligence depuis 10 mille ans
Les différences contemporaines de Q.I entre les races et entre les nations peuvent être identifiées il y a 10 mille ans déjà à partir des différences en terme de:
- capacité crânienne
- dans l’aptitude à avoir fait la transition néolithique de la chasse et de la cueillette vers l’agriculture sédentaire il y a 8000 ans
- dans le développement des premières civilisations il y a 6000 ans
- dans les avancées scientifiques, mathématiques et technologiques des 2500 dernières années
Consistency of Race Differences in Intelligence over Millennia, Richard Lynn, Personality and Individual Differences 48 (2010) 100–101.
22. Différence de fréquence des gènes de l’intelligence
- Si l’intelligence est génétique, c’est donc qu’elle est dictée par certains gènes qui ont une plus grande fréquence chez les races plus intelligentes que chez les races moins intelligentes4.3.
- De très nombreux gènes (surtout depuis les études GWAS) ont déjà été liés à un gain intellectuel et ils montrent de fait une fréquence raciale suivant l’ordre du Q.I.
Le gène de la myopie congénitale17
Le gène de la myopie congénitale: gain de 7 points de Q.I. (homozygote récessif)
- Plus haute fréquence chez les Ashkénazes (Q.I moyen de 110)
- Seconde plus haute fréquence chez les Est-Asiatiques (Q.I moyen de 105)
- Troisième plus haute fréquence chez les Européens (Q.I moyen de 100)
- Moins fréquent chez les Sud-Asiatiques/Nord-Africains (Moyen-orientaux, Q.I moyen de 84)
- Peu fréquent chez les Africains (Q.I moyen de 67)
Score polygénique (GWAS)
Une étude toute récente20 publiée dans la revue Intelligence s’est penchée sur l’ensemble des variations génétiques augmentant l’intelligence découvertes à ce jour, mises en évidence par GWAS (Genome Wide Association Study, permettant la mise en lumière de nombreux variants génétiques différents impliqués dans un trait phénotypique).
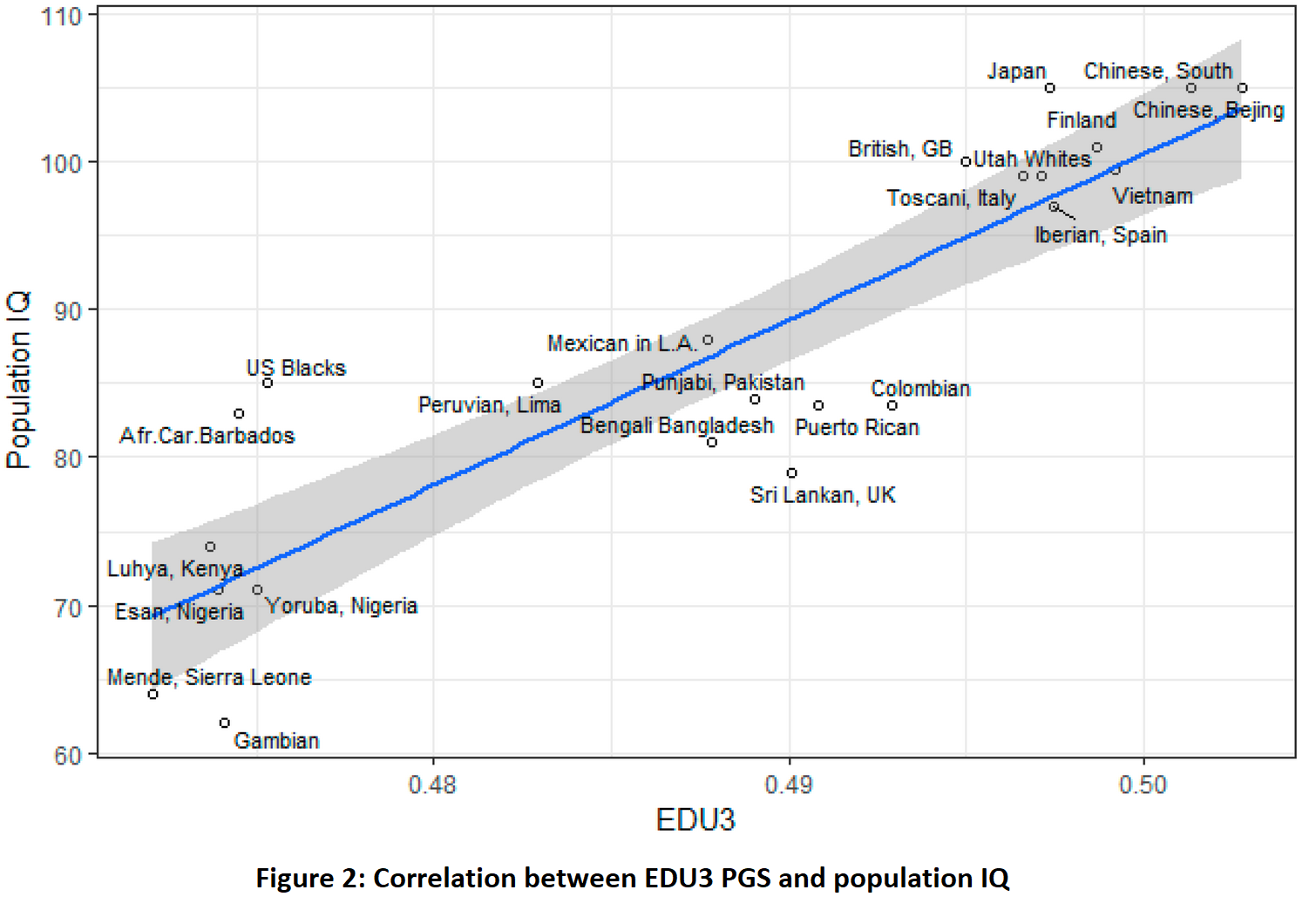
L’étude a évalué ensuite les différences raciales dans la fréquence de ces allèles, et a montré qu’elles étaient parallèles aux différences de Q.I entre populations; En d’autres termes, les races à Q.I plus élevé ont effectivement une fréquence supérieure d’allèles augmentant l’intelligence dans leur patrimoine génétique (tableau ci-dessous).

AFR pour Africains, AMR pour Amérindiens, ASN pour Asiatiques de l’est, EUR pour Européens et SAS pour Asiatiques du sud et Nord-Africains.
Si on prend en considération l’ensemble des variants génétiques augmentant l’intelligence mis en évidence à ce jour, ces allèles sont plus fréquents chez les Asiatiques de l’est (Q.I moyen de 105), suivis par les Européens (Q.I moyen de 100). Les Amérindiens (AMR, Q.I moyen de 86) et les Asiatiques du sud et Nord-africains (SAS, Q.I moyen de 84) ont une moindre fréquence et les Africains sub-sahariens ont la plus basse fréquence de ces allèles.
La fréquence de ces variants génétiques augmentant l’intelligence est donc en concordance avec l’intelligence moyenne des populations.
L’étude estime également plus précisément les fréquences de ces allèles pour différent pays et montre le lien étroit entre le Q.I moyen national et la fréquence de ces allèles dans la population.
")
Fig. 2 et Fig. 1 ci-dessus extraites de “A review of intelligence GWAS hits: Their relationship to country IQ and the issue of spatial autocorrelation” Intelligence 53 (2015) 43–50.
Ci-dessous, tableau des scores génétiques d’éducabilité (educability genetic scores) basés sur plus de 2400 variations alléliques. Ces scores sont hautement corrélés à l’intelligence, ils seraient même actuellement de meilleurs estimateurs de l’intelligence que les scores purement intellectuels, car les études réalisées ont été faites sur de plus larges échantillons (Plomin, 2018).
Comme on peut le voir, ces scores génétiques d’éducabilité sont parallèles au Q.I.
Les Est-Asiatiques ont les plus hautes fréquences d’allèles favorables, suivis par les Européens. Les Africains ont les plus basses fréquences de ces allèles.

En 2019, Dunkel, Kirkegaard et al. montrent que les Ashkénazes ont un plus haut score polygénique pour une haute intelligence.

Dunkel, Kirkegaard et al. (2019) “Polygenic scores mediate the jewish phenotypic advantage in educational attainment and cognitive ability compared with Catholics and Lutherans” Evolutionary Behavioral Sciences.
Cette étude vient corroborer les hauts scores polygéniques des ashkénazes obtenus par Piffer D. (2019)
")
Notez à nouveau comme le Q.I moyen des populations se reflète dans le score polygénique. Les populations à plus haut Q.I ont une plus haute fréquence d’allèles augmentant l’intelligence.
Conclusion
“Genome-wide association studies establish that human intelligence is highly heritable and polygenic18”
– Nature, 2011.
« Intelligence — the ability to learn, reason and solve problems — is at the forefront of behavioural genetic research. Intelligence is highly heritable and predicts important educational, occupational and health outcomes better than any other trait »
– Nature, 2018.
Références pour l’ensemble de la page
1. « Des gènes au comportement », Plomin, de Fries, Mc Clearn, Rutter, 1997.
2. Tableau inspiré de celui trouvé dans “The I.Q argument. Race, intelligence and education”, Hans Eysenck, 1971.
3. “Race differences in intelligence. An evolutionary Analysis”, Chapitre 4 pp.29-73, Richard Lynn, Washington Summit Publisher, 2006.
4. La haute stabilité des Q.I raciaux à travers le temps et quel que soit le milieu est la conséquence logique d’une haute héritabilité. Ces informations se retrouvent entre autre dans:
4.1 -“The global bell curve. Race, I.Q and inequality worldwide”, Richard Lynn, Washington Summit Publisher, 2011.
4.2 -Conférence de J.P.Rushton à American Renaissance “The Heritability of World I.Q Differences”, 2010.
4.3 -“Race differences in intelligence. An evolutionary Analysis“, Richard Lynn, Washington Summit Publisher, 2006.
5. De nombreuses études ont été effectuées pour comparer les volumes craniens dans les différentes races. Consulter “Race differences in intelligence” pour un résumé de ces études. Ce tableau récapitulatif se retrouve également dans “Le quotient intellectuel, ses déterminants et son avenir”, sous la direction de Serge Larivée, Montréal, 2009.
6. Graphique extrait de “The g factor”, 1998, Arthur R. Jensen.
7,8,9,10 et 11. “Race differences in intelligence. An evolutionary Analysis“, Richard Lynn, Washington Summit Publisher, 2006. 7. pp.64 8. pp.161 9. pp.119 10. pp.141 11. pp.105
12. “The Science of Human Diversity”, Richard Lynn, University Press Of America, 2001.
13. “Race”, John R. Baker, Oxford university press, 1974.
14. “Race, evolution and behavior”, J.P. Rushton, Charles Darwin Research Institute, 2000.
15. Rushton, J. P., & Rushton, E. W. (2004). Progressive changes in brain size and musculo-skeletal traits in seven hominoid populations. Human Evolution, 19, 173-196.
16. Sonke C.J (2000). Cross cultural differences in simple cognitive tasks:a psychophysiological
investigation. Tilberg: university press.
17. Karlsson, Jon L. (2009) Major Intelligence Gene Tied to Myopia: A Review. Mankind Quarterly, Vol. 49, No. 3-4 (Spring-Summer 2009).
18. G Davies, A Tenesa, A Payton, J Yang, S E Harris, D Liewald, X Ke, S Le Hellard, A Christoforou, M Luciano et al. (2011) Genome-wide association studies establish that human intelligence is highly heritable and polygenic. Molecular Psychiatry 16, 996-1005 (October 2011).
19. Roth B.M. (2010) « The perils of diversity. Immigration and human nature », Washington Summit Publishers.
20. Piffer D. (2015) “A review of intelligence GWAS hits: Their relationship to country IQ and the issue of spatial autocorrelation“ Intelligence 53, 43–50.
21. Plomin R. et Van Stumm S. (2018) “The new genetics of intelligence“ Nature genetics, 2018.
22. Altmann A. et al (2018) Evidence for bias of genetic ancestry in resting state functional MRI. BioRxiv.
23. Fan C.C. et al. (2015) Modeling the 3D Geometry of the Cortical Surface with Genetic Ancestry. Current Biology 25, 1-5.
24. Piffer D. (2019) « Evidence for recent polygenic selection on educational attainment and intelligence inferred from GWAS hits: a replication of previous findings using recent data »
25. Dunkel, Kirkegaard et al. (2019) « Polygenic scores mediate the jewish phenotypic advantage in educational attainment and cognitive ability compared with Catholics and Lutherans » Evolutionary Behavioral Sciences.